

特约评述
RNA疫苗的设计与递送
杨璐1,张镜明2,徐杉1,童贻刚1
(1北京化工大学生命科学与技术学院,绿色生物制造国家重点实验室,中国 北京 100029;2合肥市综合性国家中心大健康研究院,先进医药与再生医学研究所,安徽 合肥 230601)
引用本文: 杨璐, 张镜明, 徐杉, 童贻刚. RNA疫苗的设计与递送[J]. 合成生物学, 2026, 7(1): 129-151
Citation: YANG Lu, ZHANG Jingming, XU Shan, TONG Yigang. Design and delivery of RNA vaccines[J]. Synthetic Biology Journal, 2026, 7(1): 129-151
DOI: 10.12211/2096-8280.2025-085
摘 要
RNA疫苗因其快速开发和高效免疫原性成为疫苗领域的革命性技术。本文综述了RNA疫苗的设计优化与递送策略,聚焦线性mRNA、环状RNA(circular RNA,circRNA)和自扩增RNA(self-amplifying RNA,saRNA)三大类型的分子特征与应用潜力。在设计优化方面,线性mRNA通过5′帽结构、UTR(untranslated region,UTR)优化、密码子选择和poly(A)尾延长提升稳定性和翻译效率;circRNA凭借共价闭合结构抵抗核酸酶降解,实现长效表达;saRNA利用病毒复制机制扩增抗原产量,降低剂量需求。在递送系统中,脂质纳米颗粒(lipid nanoparticle,LNP)仍占据主导地位,但其可电离脂质设计和靶向配体修饰正不断优化递送效率。此外,病毒样颗粒(virus-like particle,VLP)作为新型递送载体,凭借天然的空心结构和自组装特性,兼具高生物相容性与高效mRNA装载能力,同时可模拟天然病毒入侵途径,增强细胞摄取。最后展望了其在个性化肿瘤疫苗和通用型传染病预防中的应用前景,通过整合创新递送系统(如VLP)与智能化设计,mRNA疫苗技术将迈向更精准、安全的下一代平台。
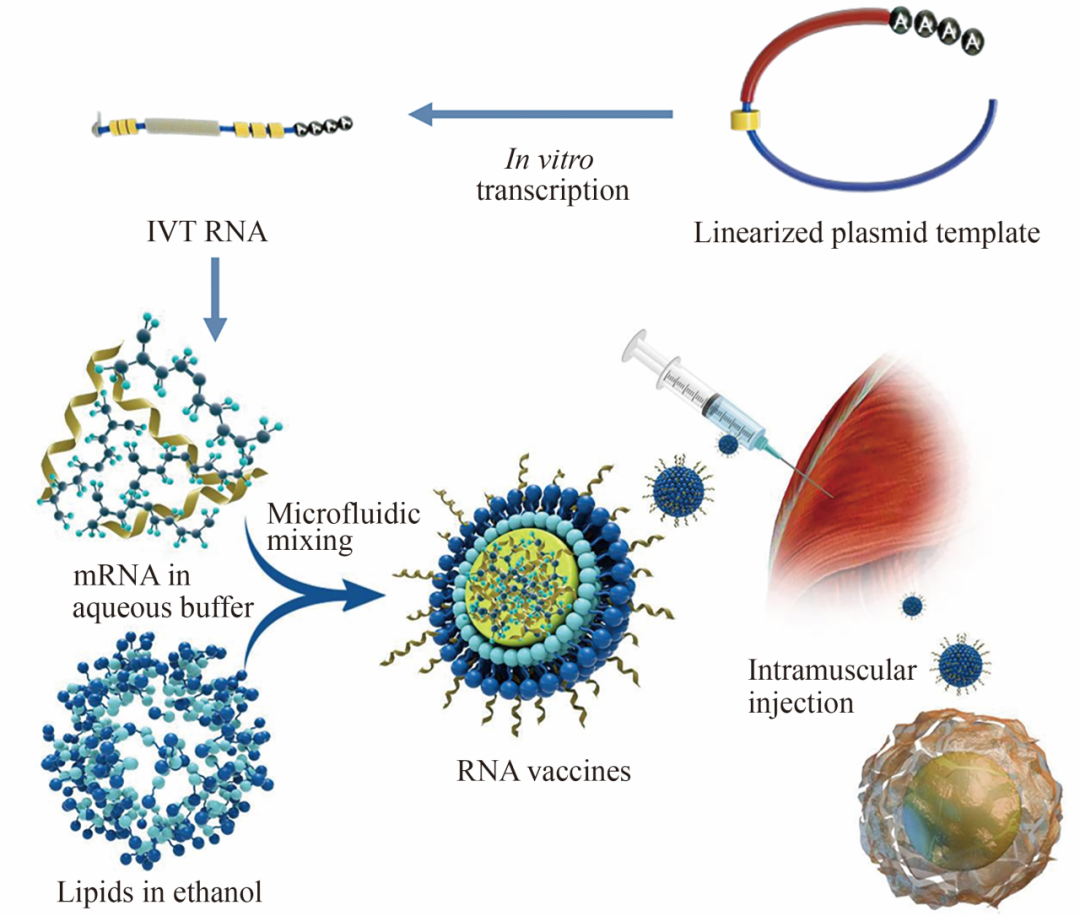
全 文
在“后疫情时代”,面对随时可能出现的突发性公共卫生事件,开发快速而普适的疫苗高效率生产平台显得尤为重要。目前获批上市的疫苗种类主要包括灭活或减毒活疫苗、重组亚单位疫苗、mRNA疫苗、活载体疫苗等。与其他类型的疫苗相比,mRNA疫苗具有易扩大生产、安全、高免疫应答等优点,完美符合快速研发的需求。
2020年以来,新冠病毒在全球范围内的肆虐推动了mRNA疫苗的迅速发展。Moderna公司研发生产的mRNA-1273以及辉瑞/BioNTech研发的mRNA疫苗BNT162b2均对新冠病毒表现出了较高的保护效力。以已获批的COVID-19疫苗为代表的线性mRNA,需要通过5′端加帽、非翻译区优化、密码子选择和poly(A)尾修饰等策略来最大化蛋白表达,同时避免先天免疫的过度激活。环状RNA凭借共价闭合结构,具有卓越的核酸酶抗性和长效表达特性,为持久抗原呈递提供了新思路。源自甲病毒基因组的saRNA则通过编码病毒复制机制实现RNA的细胞内扩增,在极低剂量下仍能维持高水平抗原表达。尽管取得这些进展,如何平衡mRNA自身免疫刺激(其本身可作为内置佐剂)与可能影响效力或引发不良反应的过度炎症仍是待解难题。
递送系统同样至关重要,因为裸露的mRNA易被降解且细胞摄取率低。LNP是目前临床主流载体,但通过对可电离脂质、聚乙二醇(polyethylene glycol,PEG)化修饰和靶向配体的持续优化,其器官靶向性和内涵体逃逸能力正在不断提升。除LNP外,VLP作为受生物启发的载体崭露头角,它兼具病毒载体的高转染效率和非复制系统的安全性。VLP能将mRNA包裹在其自组装的蛋白质外壳中,通过天然病毒入侵途径促进抗原呈递细胞(antigen-presenting cell,APC)的摄取,同时避免合成材料的细胞毒性。此外,基于聚合物的纳米颗粒和外泌体等新型平台也在探索中,以期进一步提高递送精准度和生物相容性。
本文系统综述了mRNA疫苗(线性、环状和自扩增型)的设计策略与递送技术(包括LNP、VLP等新兴系统)的最新进展,探讨如何通过mRNA分子结构与递送载体的协同优化提升疫苗效力、持久性和安全性。尽管mRNA疫苗在规模化生产与冷链稳定性方面已取得显著进展——例如通过脂质纳米颗粒(LNP)工艺优化和冻干技术提升其存储与运输适应性——但其在淋巴器官靶向递送、体内表达调控精度以及长效免疫记忆维持等方面仍面临挑战。本文进一步展望mRNA疫苗在传染病防控与肿瘤治疗领域的未来发展方向,尤其是通过智能递送系统与分子设计优化提升其临床适用性与治疗效果。通过分子生物学前沿技术与创新纳米医学的融合,mRNA疫苗的应用范畴必将远超疫情防控,开创精准医学的新纪元。
1 RNA的结构功能与设计原理
mRNA是由DNA模板转录生成的单链核糖核酸,能够携带遗传信息并指导蛋白质合成。1990年,Wolff等发现将体外转录(in vitro transcribed,IVT)mRNA或质粒DNA直接注射到小鼠骨骼肌中,可在局部诱导编码蛋白的表达,从而提出核酸编码药物的概念。基于IVT mRNA的疗法具有以下优势:①mRNA是非传染性、非整合性的,避免感染或插入突变风险,且可通过生理途径完全降解;②能够精准调控编码蛋白的表达程度和持续时间;③mRNA体外转录产量高,具有快速、廉价、可大规模生产的潜力。目前主要有三种类型的mRNA用于药物研究:非复制mRNA(non-replicating mRNA)、病毒来源的自扩增RNA(self-amplifying RNA,saRNA)与环状RNA(circular RNA,circRNA)。
1.1 非复制mRNA设计与优化
IVT mRNA是利用一个T7、T3或Sp6噬菌体RNA聚合酶从线性DNA模板体外转录产生。除了5′帽结构(5′ cap),这个线性DNA模板编码功能性mRNA的所有结构。非复制mRNA最佳结构含5′ cap、UTR、poly(A)和编码目的抗原的开放阅读框(open reading frame,ORF)[图1(a)],这种设计使其结构与真核细胞胞质中天然存在的完全成熟的mRNA分子高度相似。mRNA的稳定性和翻译由RNA/蛋白质相互作用的复杂网络控制,并取决于mRNA的一级和二级结构、它们的翻译速率以及多种降解机制。已经投入大量的努力来修饰IVT mRNA的结构元件,特别是5′ cap、5′,3′-UTR、ORF和poly(A) tail,以系统地提高其细胞内稳定性和翻译效率。这些改进最终导致在更长的时间范围内,从几分钟到一周以上,产生大量的编码蛋白。
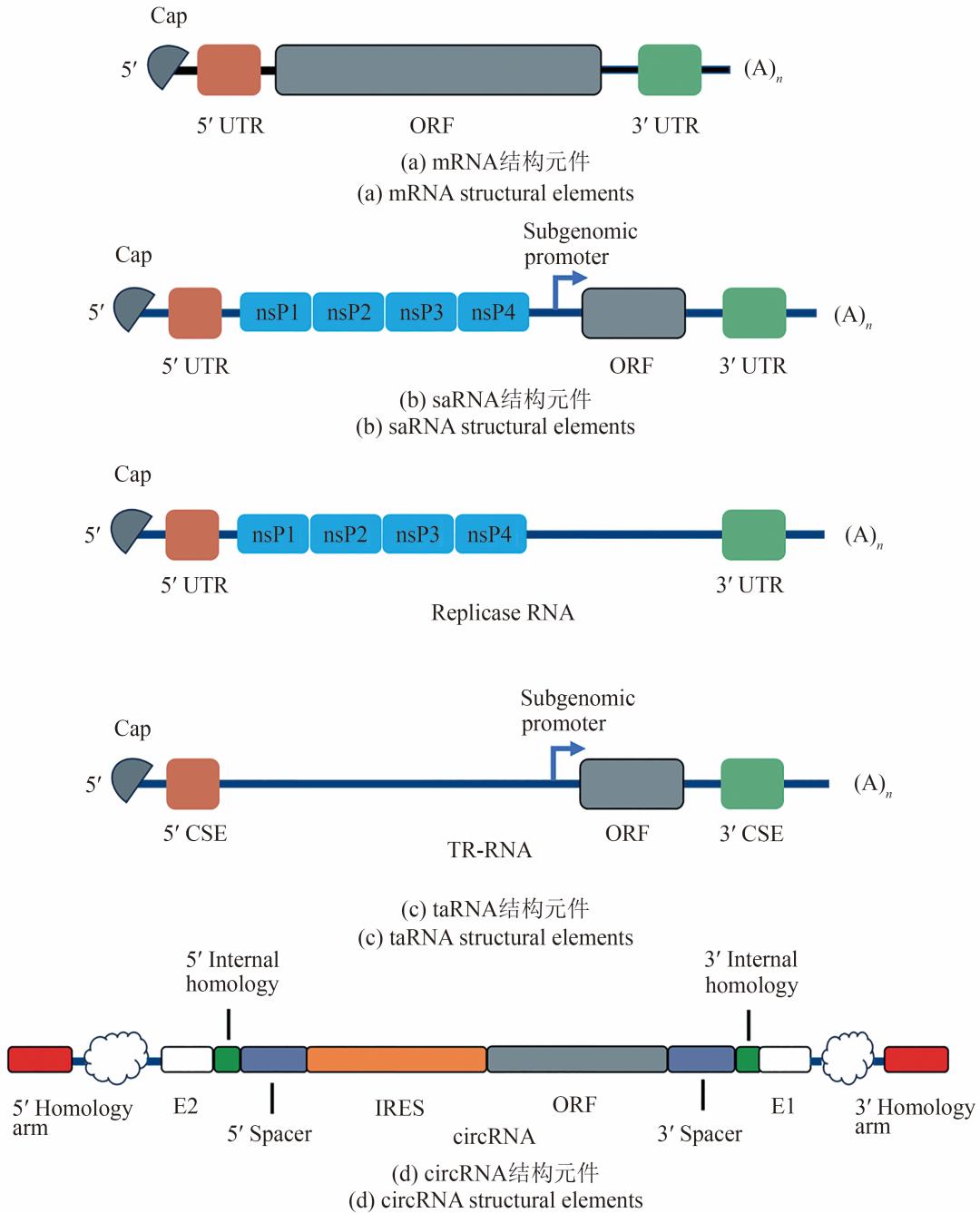
图1 RNA序列结构
1.1.1 5′ cap
5′ cap是确保mRNA高效翻译的关键元件。天然真核生物mRNA在转录过程中,通过5′,5′-三磷酸桥将7-甲基尿苷帽子连接至mRNA,形成m7 GpppN结构。体外转录(IVT)制备mRNA的过程中,常见的加帽方法包括酶法加帽和共转录加帽。酶法加帽是在线性质粒为模板进行初步合成后,通过重组的痘病毒来源的加帽酶进行第二步操作。酶法加帽的加帽率较高,生成的帽结构与最常见的天然真核生物帽结构完全相同,但其工艺相对复杂,且反应过程相较于共转录加帽引入更多的酶。共转录加帽是在体外转录反应中加入合成帽类似物,实现加帽与体外转录一步完成。然而,共转录加帽所需的帽类似物和体外转录所需的GTP核苷酸会发生竞争,导致部分mRNA未加帽,丧失翻译活性。
早期mRNA研究通过m7GpppG帽类似物(Cap0)生成的体外转录(in vitro transcribed,IVT)mRNA,即m7GpppG的m7G部分通过3′,5′-磷酸二酯键连接到RNA链的第一个核苷酸残基上。然而,两个鸟苷分子上游离的3′-OH基团会作为转录延伸的起始亲核基团,导致正向(m7G[5′]ppp[5′]G[pN]n)和反向(G[5′]ppp[5′]m7G[pN]n)两种取向产物的形成,Pasquinelli等发现1/3~1/2的帽类似物m7 GpppG反向整合进入mRNA,使得翻译机制无法识别mRNA,翻译活性下降。为解决此问题,第一代抗反向帽类似物(anti-reverse cap analogue,ARCA)被开发出来。其核心设计是对第二个鸟苷(G)核糖的3′-OH进行甲基化修饰(如m27,3′-OGpppG),从而仅保留m7G的3′-OH作为唯一的转录起始点,从结构上确保帽结构以100%正向掺入。此外,研究还发现,对第一个m7G核糖的2′-OH进行特定修饰(如m27,2′-OGpppG),即使第二个G保留游离3′-OH,也能通过立体位阻效应极高效地引导正向掺入,发挥ARCA作用。M. Mockey等发现,在小鼠树突状细胞中,编码荧光素酶的mRNA使用ARCA加帽时比m7GpppG帽类似物的表达量高约20倍。
在高等哺乳动物细胞中,Cap0结构的IVT mRNA转录本可以进一步通过转录后修饰转变为Cap1结构(m7GpppNm,其中Nm为2′-O-甲基化核苷),偶尔还会转变为Cap2结构(m7GpppNmNm)。在病毒复制过程中,免疫系统区分非自身RNA和自身RNA至关重要。第一个转录核苷中的2′-O-甲基基团在先天免疫系统中识别自身RNA并区分非自身RNA方面发挥重要作用,从而帮助抑制病毒复制和致病性。通过使用三核苷酸帽类似物,已有效解决Cap0结构的局限性。与m7GpppNmN衍生的三核苷酸帽类似物共同转录生成Cap1结构,其中第一个转录核苷具有2′-O-甲基基团。针对严重急性呼吸综合征冠状病毒(SARS-CoV-2)的三款已上市的mRNA疫苗,辉瑞-BioNTech公司的BNT162b2 mRNA疫苗通过共转录加帽Cap1;在Moderna公司的mRNA疫苗mRNA-1273中,含Cap1结构的IVT mRNA转录本是通过涉及痘病毒加帽酶和痘病毒2′-O-甲基转移酶的酶促mRNA加帽获得的;石药基团的SYS6006则是使用共转录加帽。
Cap是调控mRNA降解的关键因素之一。真核细胞中存在两类功能迥异的脱帽酶,它们在生物学功能、底物特异性及水解区域选择性方面均存在显著差异。第一种Dcp1/Dcp2复合体,是5′→3′降解途径的关键起始步骤。催化脱腺苷化mRNA的α、β磷酸键断裂,生成m7GDP和可被Xrn1降解的5′磷酸化RNA。第二种是DcpS脱帽酶,参与3′→5′mRNA降解途径。该酶能够作用于游离的m⁷GpppN二核苷酸,以及经外切体复合物通过3′端脱腺苷化和降解后释放的短链加帽mRNA,但无法切割长链RNA上的帽结构。DcpS通过区域选择性切割5′,5′-三磷酸桥的β与γ磷酸键,释放出m7GMP和下游(寡)核苷酸产物。
1.1.2 5′和3′ UTR
在5′和3′ UTR中引入特定的调控序列元件可提高IVT mRNA在细胞内翻译效率和稳定性。这些非翻译区通过与RNA结合蛋白(RNA-binding protein,RBP)的相互作用显著影响蛋白质翻译及mRNA稳定性。早期研究表明,5′ UTR的二级结构(以高GC含量为特征)会抑制mRNA翻译,因此设计IVT mRNA时,需避免使用富含GC的5′ UTR。5′ UTR应保持短小且结构松散,以便小分子核糖体与起始编码元件结合。此外,5′ UTR设计中需规避起始密码子(AUG),因其可能干扰ORF的正常翻译。近期研究表明,虽然富含腺嘌呤核苷酸(A)的5′UTR元件能实现不依赖帽子结构的翻译,但会降低未翻译mRNA的稳定性。
常见的稳定模式是3′ UTR中富含嘧啶,它们被普遍存在的α-复合物识别。α-复合物识别的最具特征的序列分别是β或α球蛋白中约180 bp或约80 bp的UTR。在一些临床前研究中,IVT mRNA的3′ UTR来自人类或非洲爪蟾β-珠蛋白基因的3′ UTR。与不含UTR的编码报告基因RNA相比,含有人β-珠蛋白3′ UTR的RNA的蛋白产量增加。Holtkamp等根据实验数据得出,使用两个人β-珠蛋白3′ UTR排列以头尾方向,进一步增强了人-珠蛋白-UTR序列的稳定作用。在终止密码子后的碱基建议为G或A,以便有效地终止翻译。此外,通过在3′UTR中加入富含AU的元件,可以实现快速的mRNA降解,从而缩短蛋白质表达时间。因此,在设计时,需减少AU元件。
1.1.3 ORF
为了优化mRNA编码目的蛋白的表达,起始密码子应位于Kozak序列中,其模式为:A/GNNATGG。核糖体将在此序列上整合N端的甲硫氨酸,并向3′端移动到下一个密码子。密码子组成会影响翻译效率。将稀有密码子替换为同义高频密码子可提高翻译产量。由于核糖体附近tRNA的氨酰化作用所致使得重复使用相同tRNA能加速翻译。相邻核苷酸和密码子也会影响翻译延伸速率和翻译效率。经过密码子优化的IVT mRNA已成功用于多项疫苗研究。另一种序列优化形式是提高GC含量,这已被证明能够在体外增加mRNA的稳态水平,并在体内提高蛋白表达,已被CureVac应用于SARS-CoV-2 mRNA疫苗候选产品CVnCoV、CV2CoV、CV0501中。此外,增加二级结构可以延长mRNA的半衰期,同时结合优化的密码子能够提高蛋白质表达。
Zhang等基于一种COVID-19 mRNA疫苗基准序列H(中国临床试验注册:CTR20210542),使用核心目标是通过优化mRNA二级结构的LinearDesign算法设计七条序列(序列A-G)。7个由LinearDesign生成的mRNA(序列A-G)的蛋白质表达水平相较于基准序列H均有显著升高。Everest Medicines(云顶新耀,HKEX 1952.HK)近日宣布,其自主研发的新型个性化治疗性mRNA癌症疫苗EVM16已正式进入临床研究阶段。该疫苗基于公司专有的AI新抗原预测算法EVER-NEO-1,通过对每位患者独特的肿瘤突变进行分析,精准筛选具有高免疫原性潜力的新抗原,并设计编码数十种肿瘤新抗原序列。采用脂质纳米颗粒(LNP)递送系统,EVM16能够高效地在体内递送新抗原编码mRNA,激活新抗原特异性肿瘤杀伤T细胞,从而抑制肿瘤生长。目前,该疫苗已在研究者发起的临床试验(IIT)EVM16CX01中完成首例患者给药,标志着这一融合人工智能与mRNA技术的个体化癌症治疗策略正式进入临床验证阶段。此外,新合生物研发的XH001注射液(受理号:CXSL2500215)作为该公司基于AI驱动的mRNA技术平台开发的首款个性化新抗原疫苗,通过其自主研发的NeoCura® AI ALPINE系统,整合患者个体肿瘤突变谱与HLA分型数据,高效筛选具有高免疫原性的新抗原,进而构建针对个体定制的mRNA疫苗制剂。该疫苗通过编码特异性新抗原,激活机体肿瘤特异性T细胞免疫应答,靶向清除残余肿瘤细胞,从而从源头上降低肿瘤复发风险。目前已开展的研究者发起临床研究(IIT)初步数据显示,XH001安全性及耐受性良好,并可有效诱导特异性T细胞免疫反应,为其后续Ⅰ期临床试验的开展奠定了坚实基础。
然而,在某些情况下可能需要避免使用优化密码子。某些蛋白质需要依赖稀有密码子实现的慢速翻译来确保正确折叠。对于某些体外转录mRNA编码的疫苗,维持原始开放阅读框(ORF)也可能更为有利。当体外转录mRNA因核糖体移码以不同阅读框翻译时,或当翻译从内部或CUG起始密码子启动时,可能会产生强效隐性T细胞表位。而密码子优化应会消除这些抗原肽的重要来源。
1.1.4 poly(A)尾
poly(A)尾通过与5′端帽结构、内部核糖体进入位点(internal ribosome entry site,IRES)及其他多种调控元件协同作用,共同调节mRNA的稳定性和翻译效率。Sachs和Davis提出poly(A)尾通过PAB蛋白介导与60S核糖体亚基之间的相互作用对调控80S翻译起始复合体的形成具有重要作用。由于它们的双重调节作用,通过添加5′ cap或poly(A)尾,mRNA翻译效率和稳定性都得到了提高。在烟草原生质体中,未加帽的、含poly(A)尾的Luc mRNA比未加帽的、不含poly(A)尾的Luc mRNA的荧光素酶表达量和半衰期分别高1.5倍和1.4倍。此外,poly(A)尾是加帽功能最优化的必要条件。当poly(A)尾添加到加帽形式的mRNA时,含poly(A)尾的mRNA表达量是不含poly(A)尾mRNA的21倍。在DC中的分析表明,poly(A)尾的最佳长度在120~150个核苷酸之间。在 BioNTech公司公开的专利中,含有120 nt的poly(A)尾的mRNA的稳定性和翻译效率高于poly(A)尾长度为16 nt、42 nt、51 nt和67 nt 的mRNA。Pfizer和BioNTech联合研发并已上市的mRNA疫苗BNT162b2,poly(A)尾长度为110个核苷酸。该poly(A)尾,包括一段由30个腺嘌呤残基组成的序列,接着是一个10个核苷酸的连接序列,再加上另一段由70个腺嘌呤残基组成的序列。在Karekar等提出的RNA-LNP疫苗开发的标准化操作流程中,同样使用该方法。而Moderna 公司的mRNA-1273疫苗未说明具体的poly(A)尾使用策略。CureVac公司的 mRNA疫苗CVnCoV的poly(A)尾,则是使用由64个腺嘌呤残基组成的序列。IVT mRNA可通过模板载体编码获得poly(A)尾,也可以在转录后通过重组poly(A)聚合酶单独反应添加poly(A)尾。然而,酶促加poly(A)尾的方法制备的RNA都会包含具有不同长度poly(A)尾,这在临床应用中不被美国FDA认为可接受。
1.2 saRNA的结构与优化
saRNA疫苗的基础源于正链单链RNA[(+)ssRNA]病毒(例如甲病毒、黄病毒、慢病毒、麻疹病毒和弹状病毒)的自我复制特性。这些单链RNA病毒通过其编码的非结构蛋白(nonstructural proteins,nsP)基因,形成以RNA依赖型RNA聚合酶(RNA-dependent RNA polymerase,RdRp)复合体为核心的病毒复制机制——该复合体通过与宿主因子的特异性相互作用,驱动病毒基因组的复制与表达。saRNA疫苗理论上能够进行多轮RNA复制,从而在体内产生高产量的疫苗抗原,降低疫苗剂量,从而实现更安全的免疫接种。
1.2.1 saRNA结构原理
saRNA虽然长度显著大于非复制型mRNA(约9~10 kb),但仍保留了mRNA的基本结构元件[5′ cap、5′ UTR、3′ UTR以及长度可变的poly(A)尾][图1(b)]。目前研究最为深入的saRNA分子源自α病毒(如委内瑞拉马脑炎病毒(Venezuelan equine encephalitis virus,VEEV)、辛德毕斯病毒(Sindbis virus,SINV)和塞姆利基森林病毒(Semliki forest virus,SFV))的基因组结构。在构建saRNA疫苗时,研究人员用抗原基因替换α病毒结构蛋白编码区。体外转录生成saRNA的质粒模板包括:①一个指定的噬菌体编码的DNA依赖型RNA聚合酶(DNA-dependent RNA polymerase,DdRP)启动子(如T7、T3或SP6);②一个来源于甲病毒的5′-UTR;③编码甲病毒nsP(nsP1、nsP2、nsP3和nsP4)的基因,以产生活性RdRp;④病毒的26S亚基基因启动子序列;⑤编码疫苗抗原的ORF;⑥一个来源于甲病毒的3′-UTR;⑦一个poly(A)尾;⑧一个位于3′-末端的独特限制酶切位点。
saRNA疫苗相较于传统mRNA疫苗具有多重优势,其核心在于能在宿主细胞内自主复制,这种特性既增强了免疫原性又大幅降低了初始剂量需求。Vogel等证实,在小鼠流感模型中,传统mRNA疫苗需要80 µg剂量才能诱导保护性免疫反应,而saRNA疫苗仅需1.25 µg即可达到同等保护效果。McKay团队针对SARS-CoV-2的最新研究进一步凸显了saRNA疫苗的剂量优势——编码预融合稳定刺突蛋白的saRNA疫苗在临床前模型中,仅用1 µg剂量就成功诱导强效中和抗体反应并实现免疫保护。针对SARS-CoV-2研发的saRNA疫苗VLPCOV-01,Ⅰ期临床试验数据显示:已完成两剂BNT162b2疫苗接种的受试者随机接受0.3 μg、1.0 μg或3.0 μg VLPCOV-01、30 μg BNT162b2或安慰剂的单次肌内注射后,0.3 μg VLPCOV-01组的几何平均滴度是30 μg BNT162b2组的1.59倍。
1.2.2 反式自扩增RNA(trans-amplifying(ta)RNA)结构原理
saRNA疫苗进一步发展出taRNA疫苗技术。该体系采用双RNA组分[图1(c)]:第一种是体外转录的mRNA,编码甲病毒复制酶,可在体内直接翻译;第二种为反式复制子(trans-replicon,TR)RNA,编码相应的抗原,由亚基因组启动子(subgenomic promoter,SGP)调控。由于TR-RNA含有甲病毒5′和3′保守序列元件(conserved sequence element,CSE),甲病毒复制酶可对其进行反式扩增。最初Blakney等基于VEEV骨架,使用荧光报告基因作为抗原,建立了名为“splitzicon”的分割复制子系统,明确了taRNA自扩增所需组分。Beissert与其团队构建的针对流感病毒的taRNA候选疫苗包含非复制型复制酶mRNA(non-replicative RNA replicon,nrRNA-REPL)和表达流感病毒血凝素的TR-RNA,与saRNA疫苗相比,该疫苗能以更少抗原RNA剂量诱导保护性免疫应答。此外,针对流感等季节性疫苗,可将nrRNA-REPL作为不变组分提前大规模生产储备,而仅需针对可变的、高效抗原编码RNA进行按需的新工艺开发和生产。基于基孔肯雅病毒(Chikungunya virus,CHIKV)的taRNA疫苗候选株,采用初免-加强免疫策略,仅需1.5 μg(1.25 μg 复制酶RNA和0.25 μg TR-RNA)RNA剂量即可诱导强效体液与细胞免疫,成功保护小鼠抵御106 PFU病毒攻击。Schmidt等设计一种新型双价taRNA疫苗策略,通过递送三种RNA(一种编码CHIKV复制酶,两种分别编码CHIKV和RRV包膜蛋白的TR-RNA)实现多抗原免疫。
1.2.3 局限性和优化策略
基于甲病毒的saRNA疫苗,存在重组风险隐患,特别是存在共感染病毒的情况下。理论上,细胞内甲病毒复制子来源的saRNA可能与病毒RNA发生重组,从而产生新型或变异的病毒株。VEEV因其复制效率远高于其他甲病毒,是目前saRNA设计的常用平台,但VEEV的高复制率同时会提升突变和重组事件的发生概率,进而增加嵌合病毒形成的风险。Hick等指出saRNA的自扩增特性(特别是采用VEEV等高复制病毒骨架时),在病毒共感染或潜伏病毒基因组存在的环境中,可能引发非预期的基因漂变。甲病毒间的自然重组已有历史证据,且被证实与新发病毒的出现相关。西方马脑炎病毒(Western Equine encephalitis virus,WEEV)就是SINV与VEEV群自然重组产生的。
针对这一问题,可采用taRNA系统作为替代方案。虽然这种分离设计本身并不能降低与野生型病毒的重组概率,但由于复制机制不具备自主增殖能力,从而减少了重组可能性,可有效控制风险。Beissert等的研究证实,该策略在保持免疫原性的同时减少抗原编码复制子的大小。然而,该系统需要同时制备两种RNA组分,且目前尚未开发出高效的体内递送方式。
1.3 circRNA的设计与合成
circRNA是一类具有共价闭合环状结构的单链RNA分子。随着生物技术和分子医学的发展,人工设计的circRNA已被开发为一类用于疾病治疗和预防的新型疫苗。与线性mRNA疫苗相比,circRNA具有多种优势:①由于其具有特殊的环状结构,circRNA无需任何修饰即可表现出高度稳定性和抗RNase降解性,易于储存;②相较于线性mRNA需要使用修饰核苷酸降低免疫原性,未经修饰的circRNA具有更低的免疫原性和更低的细胞毒性;③circRNA具有延长抗原产生成效和持久免疫应答的能力。
1.3.1 circRNA的结构
基于重组的Ⅰ型催化内含子系统已被用于体外环化多种短RNA序列(58~124 nt)中,其环化效率可达90%。Chen等采用具有自催化剪接功能的噬菌体T4胸苷酸合成酶(thymidylate synthase,td)基因的Ⅰ型内含子进行体外circRNA合成。用于IVT的DNA模板序列包含3′内含子片段、绿色荧光蛋白(green fluorescent protein,GFP)和脑心肌炎病毒(encephalomyocarditis virus,EMCV)的IRES序列,以及5′内含子片段。体外转录后,按预期生成1.5 kb的circRNA。Wesselhoeft等工程化Ⅰ型催化内含子系统,能够实现长度达5 kb的序列环化,并且针对不同序列组成的编码区,环化效率接近100%。用于环化的前体RNA包含5′同源臂、3′内含子片段、外显子片段2、5′内部同源序列、5′间隔区、EMCV IRES、编码区域、3′间隔区、3′内部同源序列、外显子片段1、5′内含子片段、3′同源臂[图1(d)]。优化后的circRNA不仅能高效表达大量蛋白,还可通过HPLC有效纯化。2022年,北京大学魏文胜教授团队利用工程化Ⅰ型催化内含子系统,编码区插入源自人类组织型纤溶酶原激活剂(tissue plasminogen activator,tPA)的信号肽序列、RBD序列以及来源于噬菌体T4纤维蛋白的三聚化结构域,开发了针对SARS-COV-2的环状RNA疫苗。该研究开展了小鼠及恒河猴的动物实验,环状RNA疫苗表现出比线性mRNA疫苗更为持久和稳定的抗原表达。Jaffrey团队采用Tornado circRNA表达系统,开发了一种在哺乳动物细胞中合成并高效表达环状mRNA的新方法。该团队构建的用于表达编码SARS-CoV-2刺突蛋白的Tornado载体,包含CMV启动子、5′端Twister核酶(包含环化连接序列)、柯萨奇病毒B3型(Coxsackie virus B3,CVB3) IRES、S蛋白、3′端Twister核酶(包含环化连接序列)、终止子。将此载体转染HEK293T细胞,可实现全长4719个核苷酸的环化。
1.3.2 环状RNA的合成
通常通过IVT方法,利用T7或SP6等噬菌体RNA聚合酶,以DNA为模板大量扩增目标序列生成线性前体。前体线性RNA通过共价闭合形成环状结构获得circRNA,该过程通常由化学、酶促或核酶连接方法介导。
(1)化学法
1988年提出的核酸链化学连接法利用DNA链的5′磷酸基团与3′羟基,在2-(N-吗啉代)-乙烷磺酸(MES)催化下,通过BrCN生成的氰酸酯中间体形成磷酸二酯键。该方法同样适用于短链环状RNA的合成,但需借助寡核苷酸“夹板”拉近5′和3′端。
1999年,Micura等开发了一种从头合成环状RNA的方法。该方法通过2′-O-TOM RNA磷酰胺实现RNA链全自动组装,利用标准磷酸三酯化学在固相支持物上合成环状RNA,并通过选择性切割获得高纯度的产物,环状RNA的平均产率为15%。
(2)酶法合成
酶促合成法主要利用T4 RNA连接酶Ⅰ和Ⅱ,在ATP依赖的条件下催化线性RNA前体的5′-磷酸基与3′-羟基形成3′,5′-磷酸二酯键。
T4 RNA连接酶Ⅰ在1972年被用于线性前体的环化。然而,该方法可能产生分子间连接的聚体,但通过优化反应条件,环化产物可成为主产物。最短6~8 nt的RNA链即可环化。T4 RNA连接酶Ⅰ的对连接位点的供体和受体核苷酸的偏好不同:3′端核苷酸受体的偏好性为A>G≥C>U,5′端核苷酸受体为pC>pU>pA>pG。其环化效率受线性RNA前体的二级结构影响,使用辅助脱氧寡核苷酸可以防止线性RNA折叠成不利结构,从而提高环化效率。在线性辅助寡核苷酸的存在下,环状RNA的产率为40%,发夹辅助环状RNA的产率接近100%。除了通过辅助连接寡核苷酸外,还可以使用夹板使活性端靠近在一起,使供体和受体RNA的末端保持2~3个核苷酸单链。此外,通过设计线性RNA前体的5′和3′末端为单链区域,利用内部二级结构拉近两端也可促进环化。
2002年,Ho等描述了T4 RNA连接酶Ⅱ的分子间和分子内连接活性。与T4 RNA连接酶Ⅰ相比,T4 RNA连接酶Ⅱ能够更有效地连接dsRNA底物上的缺口,而不是连接ssRNA的末端。根据T4 RNA连接酶Ⅱ的这一特性,在设计线性RNA前体序列时,5′供体和3′受体端需要通过内部二级结构保持较近的空间距离。在噬菌体KVP40中存在一种与T4 RNA连接酶Ⅱ同源的蛋白。它以ATP依赖的方式催化单链小RNA的环化。此外,T4 RNA连接酶Ⅱ存在截短版本(249个氨基酸),可在无ATP条件下催化预腺苷化的RNA或DNA与RNA的连接,用于RNA环化。这些特性使T4 RNA连接酶在RNA环化中具有广泛应用潜力。
(3)核酶催化合成
与只能在体外应用的化学和酶连接方法不同,一种基于Ⅰ型内含子自剪接原理开发的环化方法,被称为PIE策略,可以在体外和体内产生环状RNA。利用Ⅰ型内含子的自剪接特性的方法应用较为广泛,更适用于长链RNA。且相比于化学法和酶法合成环状RNA需要在体系中引入额外的化学和生物催化剂,核酶法只需要在体系中添加GTP和Mg2+作为辅助因子。该方法的原理为Ⅰ型内含子作为核酶催化的RNA自剪接,该过程包括两步转酯反应。首先,一个外源的鸟苷(G)结合到内含子催化中心的G结合位点,其3′-OH攻击5′剪接位点的磷酸二酯键,与内含子的5′端第一个核苷酸形成3′,5′-磷酸二酯键,并释放出5′-外显子。随后,游离5′-外显子的3′-OH攻击3′剪接位点的磷酸二酯键,使5′-外显子与3′-外显子发生连接并释放内含子。基于该机制,M. Puttaraju等于1992年发明了Ⅰ型内含子-外显子置换系统(permuted intron-exon,PIE),利用核酶的自剪接特性形成一个外显子-外显子共价连接的环状RNA。当本体的5′端剪接位点位于本体剪接位点的3′端处时,RNA剪接反应可产生环状外显子序列。在这种情况下,外源核苷酸序列插入外显子和内含子位点之间,产生包含外源RNA片段的环状RNA。在此PIE系统基础上,Wesselhoeft等做出了一系列优化,增加同源臂和间隔序列产生更大的环状RNA,增加IRES增加蛋白翻译效率。PIE策略在其被提出之后的三十余年中得到了广泛的应用。
此外,除了Ⅰ型内含子外,Ⅱ组内含子也可以用同样的重排方法用于体外合成环状RNA,其机制与Ⅰ型内含子相似。Mikheeva等证明,可以将酵母线粒体基因组中Ⅱ型内含子重排,构建 PIE结构。与Ⅰ型内含子PIE系统不同,Ⅱ型内含子的使用导致剪接机制不需要原生外显子,从而避免引入外源序列。Sun等构建基于ⅡC型自剪接内含子针对RSV的新型环状RNA疫苗,在体内可有效激发以Th1型偏向为主的细胞和体液免疫应答。
1.3.3 环状RNA纯化
PIE策略合成的环状RNA中主要存在的杂质包括未成功环化的线性RNA前体、剪接出的RNA内含子片段、核苷三磷酸以及开环RNA。由于线性RNA前体和开环RNA的分子量与对应circRNA相近,去除线性RNA杂质成为circRNA疫苗生产中的难点。可采用HPLC或RNase R酶切法去除线性RNA前体等杂质。
环状RNA由于其共价闭合的环状结构,可以在一定程度上抵抗核酸外切酶的降解,因此可以利用该特点对环状RNA进行纯化。RNase R能消化线性RNA,但不能消化环状RNA、套索RNA、3′突出末端少于7个核苷酸的双链RNA以及具有复杂二级结构的tRNA、5S RNA等。RNase R酶切线性RNA前体的主要问题在于其复杂的二级结构会降低对RNase R的敏感性。尽管高温加热和化学试剂可以破坏二级结构,但这些方法可能增加后续纯化步骤的难度。由于线性RNA在3′端为单链状态时更易被RNase R降解,可通过引入无二级结构的元件[如poly(A)尾]来提高降解效率。然而,低剂量RNase R可能无法充分降解线性RNA,而高剂量又可能导致circRNA断裂,因此需要优化反应条件。此外,RNase R价格昂贵,限制了其工业应用。
HPLC是一种灵敏度高、精密度高、适用范围广的色谱技术,目前广泛应用于环状RNA的纯化,其中体积排除色谱(SEC-HPLC)是最常用的填料。通过结合RNase R酶切,SEC-HPLC能够分离环状RNA、开环环状RNA和线性RNA前体,并显著提高环状RNA纯度,降低其免疫原性。然而,由于这些RNA分子具有相近的分子量和二级结构,色谱图中仍可能出现重叠峰,同时环状RNA在纯化过程中容易自发开环。现有方法尚需优化,可尝试离子交换色谱或反相色谱与SEC联用,同时改进流动相条件(如调节pH值、加入柠檬酸钠)以进一步提高纯化效率并减少环状RNA的开环风险。
1.4 免疫原性调节
外源性mRNA本质上具有免疫刺激性,因为它会被多种细胞表面、内体和细胞质中的先天免疫受体识别。酶促合成的mRNA制备中含有双链RNA(double-stranded RNA,dsRNA)污染物,这是IVT反应的异常产物。作为病毒基因组和复制中间体的模拟物,dsRNA是一种强效的病原相关分子模式(pathogen-associated molecular patterns,PAMP),能够被多个细胞区室中的模式识别受体感知。含有dsRNA污染物的IVT mRNA被识别后,会导致强效的Ⅰ型干扰素产生,从而上调蛋白激酶R(protein kinase R,PKR,也称为EIF2AK2)和2′,5′-寡腺苷酸合成酶(2′,5′-oligoadenylate synthetase,OAS)的表达和激活,分别引发翻译的抑制以及细胞mRNA和核糖体RNA的降解。Karikó团队研究证实,通过反相快速蛋白液相色谱(reversed-phase FPLC,RP-FPLC)或高效液相色谱(high performance liquid chromatography,HPLC)等色谱方法可有效去除体外转录mRNA中的dsRNA杂质,其中FPLC纯化能使mRNA在人类原代树突细胞中的蛋白表达量提升高达1000倍。此外,单链mRNA分子本身在外源性传递至细胞时也是一种PAMP。单链寡核糖核苷酸及其降解产物会被内体传感器TLR7和TLR8检测到,从而引发Ⅰ型干扰素的产生。
通过整合修饰的核苷酸[假尿苷(Ψ)、1-甲基假尿苷(m1Ψ)等],不仅可以防止TLR7、TLR8以及其他先天免疫传感器的激活,还能够部分抑制对dsRNA类型的识别,增强翻译能力。Karikó等将多组编码海肾荧光素酶的mRNA与lipofectin复合后转染293细胞,结果表明含Ψ修饰的mRNA翻译效率约为未修饰mRNA的10倍,而含m5C修饰的mRNA则产生4倍增强效果。
三大mRNA疫苗巨头公司Moderna、BioNTech和Curevac研发的mRNA-1273、BNT162b2和CVnCoV疫苗,其保护率分别为94.1%、95%和48%。CVnCoV的有效性未能达到世界卫生组织(WHO)的基准要求,已终止研发。分析三款mRNA疫苗的设计,不含5′ UTR和修饰核苷酸可能是导致CVnCoV疫苗保护率较低的原因。
2 mRNA疫苗递送策略
高效的体内mRNA递送是实现治疗的关键。外源mRNA必须穿透脂质膜屏障才能进入细胞质,进而翻译为功能性蛋白(图2)。多种细胞类型可自发摄取裸露mRNA。通过清道夫受体介导的内吞作用进入细胞的裸露mRNA会积聚在溶酶体中,仅少量mRNA能泄漏至细胞质。mRNA释放至细胞质的机制尚未明确表征,且可能因细胞类型而异。此外,mRNA还能通过外泌体在细胞间进行交换。对大多数细胞而言,mRNA的主动摄取效率低下,且在低剂量时即达饱和。未成熟树突状细胞(DC)是个例外——这些细胞在环境采样过程中特化于持续吞噬细胞外液,能通过巨胞饮作用以线性非饱和方式高效摄取mRNA,其浓度适应范围可达数个数量级。因此,要向大多数细胞类型递送mRNA,需要合适的制剂保护体外转录(IVT)mRNA免受细胞外RNase介导的降解,并促进其进入细胞。目前mRNA疫苗递送主要有两种基本策略:第一种是体外将mRNA导入树突状细胞(dendritic cell,DC)后将转染的细胞重新输注;第二种是直接通过注射途径递送携带或不携带载体的mRNA。体外DC转染能精确控制靶细胞类型、转染效率等参数,但作为细胞疗法,这种疫苗接种方式成本高昂且操作复杂。相比之下,mRNA直接注射更为快速经济。
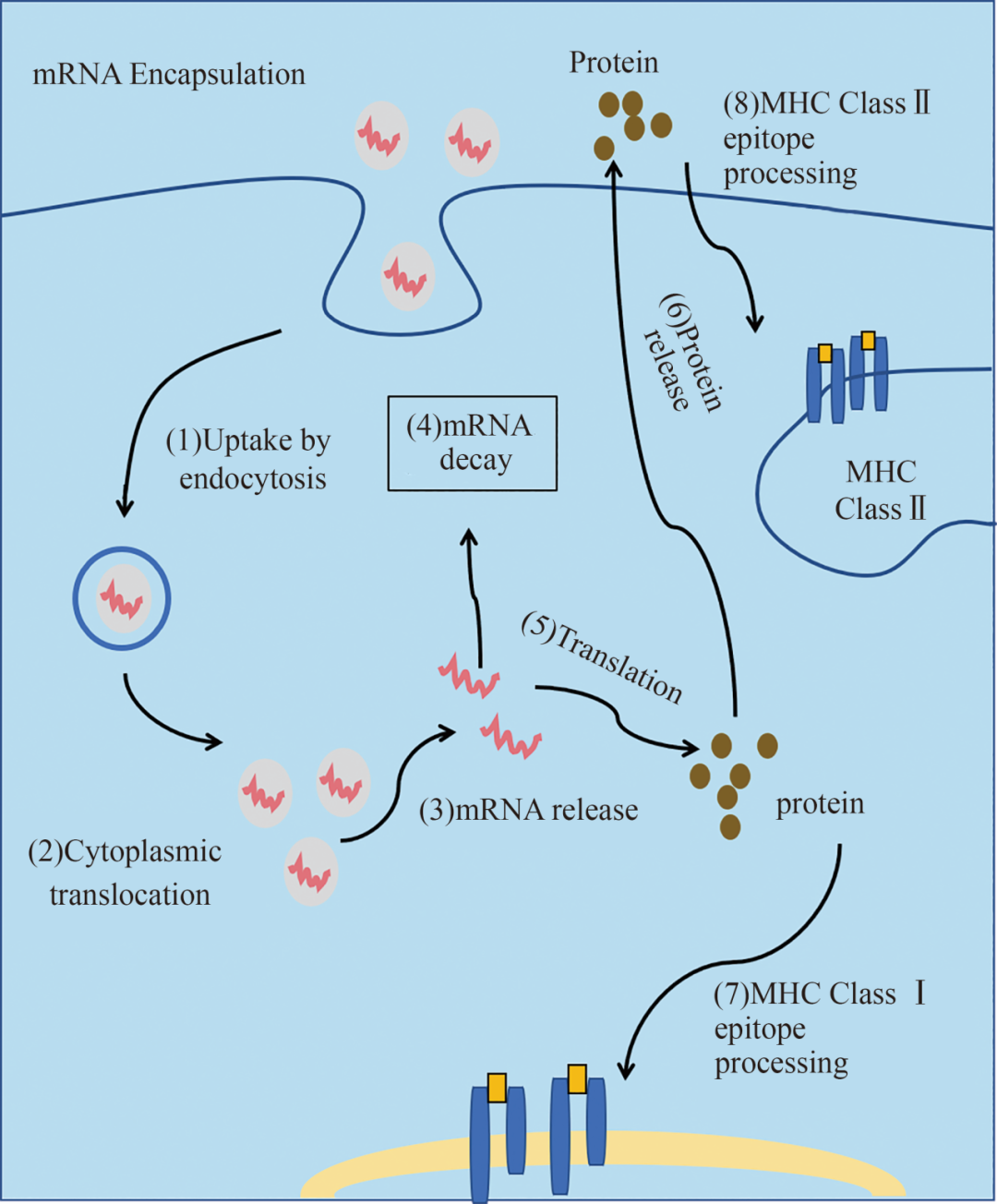
图2 mRNA递送原理
[(1)包封后的mRNA被细胞摄取内吞,进入内体途径;(2)包封后的mRNA释放至细胞质;(3)mRNA从递送载体中释放;(4)翻译终止伴随mRNA降解,由核酸外切酶催化;(5)mRNA翻译成蛋白质;(6)蛋白质通过自分泌、旁分泌或内分泌机制分泌至胞外行使功能;(7)蛋白质产物需被降解为抗原肽表位,这些表位被装载至主要组织相容性复合体(MHC)分子上,呈递至细胞表面供免疫效应细胞识别;(8)在抗原呈递细胞中,为获得T细胞辅助以增强免疫应答持续性,需将蛋白质产物导向MHCⅡ类分子装载区。]
2.1 体外负载DC法
DC作为连接先天免疫与适应性免疫的关键桥梁,是调控T细胞应答的主要抗原呈递细胞。其通过内吞和蛋白水解加工抗原,并分别通过主要组织相容性复合体(major histocompatibility complex,MHC)Ⅰ类和Ⅱ类将抗原呈递给CD8+和CD4+T细胞,从而启动适应性免疫应答。此外,DC还可将完整抗原呈递给B细胞以激发抗体反应。DC对mRNA转染也具有高度适应性,这些特性使其成为mRNA疫苗极具吸引力的靶点。
尽管研究表明DC能通过多种内吞途径摄取裸露mRNA,但体外转染通常采用电穿孔技术以提高效率——该方法通过高压脉冲形成细胞膜孔隙,使mRNA分子直接进入细胞质。这种递送方式因无需载体分子即可实现高效转染而备受青睐。经体外负载mRNA的DC随后被重新输注到自体疫苗接种者体内以启动免疫反应。大多数体外加载的DC疫苗主要诱导细胞介导的免疫反应,因此主要用于癌症治疗。
2.2 基于脂质递送系统
在非病毒核酸载体中,基于脂质递送系统(如LNP、脂质体、脂质复合物)已成为最常用的递送平台(图3)。DOTMA(N-[1-(2,3-二油酰氧基)丙基]-N,N,N-三甲基氯化铵)是首个用于体外递送编码荧光素酶IVT-mRNA的合成阳离子脂质,已成功实现了多种(如人、大鼠、小鼠、非洲爪蟾和果蝇)细胞的体外转染。研究发现DOTMA衍生物DOTAP(1,2-二油酰基-3-三甲基铵-丙烷)具有较低的制备成本和更高的递送效率。为提升DOTAP介导的IVT-mRNA转染效率,Zohra等在DOTAP脂质体载体中引入碳酸盐磷灰石(一种对核酸具有强亲和力的无机晶体),使转染效率较单纯DOTAP对照组提高5~15倍。后续研究发现,在DOTAP-磷灰石颗粒中整合纤维连接蛋白(靶向α5β1整合素的识别基序),使HeLa细胞中IVT-mRNA的转基因表达(定性和定量)均优于非靶向颗粒。后续研究发现两性离子脂质DOPE(1,2-二油酰-sn-甘油-3-磷酸乙醇胺)与DOTAP联用可显著增强基因表达。阳离子脂质还能用于制备阳离子纳米乳(CNE),通过均质化含辅料的水相/油相后与mRNA复合而成。Bogers等研究发现使用CNE递送编码HIV C亚型包膜糖蛋白的saRNA疫苗,在恒河猴中诱导了强效免疫应答。Luisi等利用sa mRNA技术平台结合阳离子纳米乳(cationic nanoemulsion,CNE)递送系统开发了针对寨卡病毒(Zika virus,ZIKV)现用现配的候选疫苗。其中,VRC5283 SAM(CNE)疫苗在小鼠和非人灵长类攻毒模型中均能持续诱导针对ZIKV的完全免疫保护。
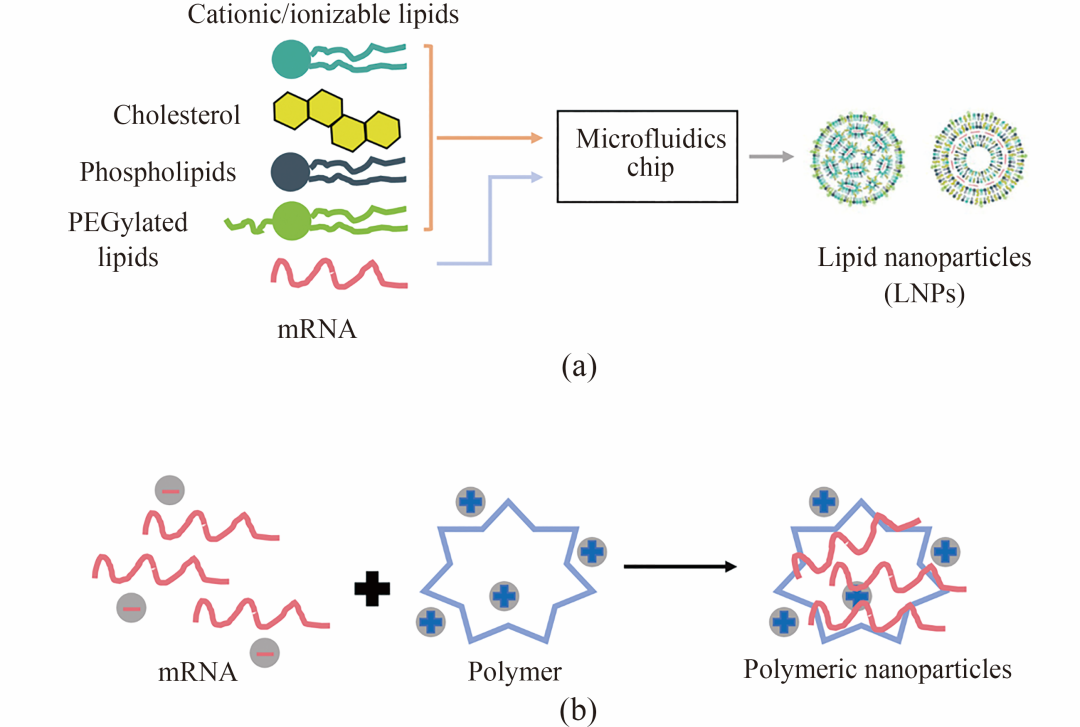
图3 递送系统的常见结构
[(a) 脂质纳米颗粒:通常通过微流控技术制备,成分包括传统(阳离子或可离子化)脂质、功能化脂质、胆固醇及辅助脂质。(b) 聚合物载体:包括聚乙烯亚胺(PEI)、可离子化两亲性Janus树枝状大分子(IAJD)和壳聚糖等,可通过简单混合包裹mRNA形成稳定的纳米颗粒。]
随着微流控技术的发展,现可采用阳离子/可电离脂质、电中性磷脂、胆固醇和PEG化脂质高效制备LNP,大幅提升mRNA的包封率和转染效率。美国FDA批准的Patisiran、BNT162b2和mRNA-1273三款药物均基于LNP技术。其各组分功能明确:阳离子/可电离脂质通过正电荷结合mRNA并协助内体逃逸;磷脂(如DOPE、DSPC)和胆固醇维持结构稳定;PEG化脂质(如DMG-PEG2000)可减少血浆蛋白非特异性结合,延长体内半衰期。在临床应用中,LNP技术具有技术成熟、递送效率高和模块化设计等优势,已有多款产品获批且生产工艺易于放大;然而仍面临靶向性局限、潜在毒性和长期安全性未明等挑战,且多数制剂依赖低温储存,对物流要求较高。
当前LNP研发聚焦三大方向:新型脂质发现、组分优化和功能化修饰。传统设计策略通过调控氨基头基、疏水尾链和连接桥来改变脂质特性。功能化脂质方面,Oyama等研究开发的维生素E支架脂质(ssPalmE)能激活Ⅰ型干扰素信号通路增强抗肿瘤免疫;Tang等报道的苯硼酸修饰阳离子脂质PBA-BADP可与癌细胞表面过表达的唾液酸特异性结合,提升纳米颗粒内化效率。针对安全性问题,Bang团队设计的海藻糖糖脂TDO通过氢键稳定颗粒结构,在保持递送效能的同时降低可电离脂质毒性,体现了LNP技术向高效化与安全化并重的发展趋势。值得注意的是,冻干疫苗技术的突破为LNP制剂提供了全新的稳定性解决方案——通过冷冻干燥工艺可实现mRNA-LNP疫苗在25 ℃条件下长期储存,且不影响其生物活性与免疫原性,这标志着LNP技术正朝着高效化、安全化与制剂稳定化并重的方向发展。
2.3 基于聚合物递送系统
带负电的IVT-mRNA可通过静电作用自发与阳离子聚合物形成复合物。PEI是目前研究最广泛的核酸递送聚合物,其丰富的氨基可提供高密度正电荷以实现mRNA封装,并通过“质子海绵”效应实现出色的内体逃逸。然而PEI介导的mRNA递送存在效率-毒性失衡问题:高分子量PEI虽递送效率优异但细胞毒性显著,低分子量PEI毒性较低却效率不足。Guo等基于低分子量PEI(1.8 k)构建了14种聚合物候选库,最终筛选出性能最优的含有八碳烷基链的POctS。POctS不仅在mRNA递送中展现出显著的部位特异性,还可直接与STING蛋白相互作用,诱导关键信号分子TBK1和IRF3的磷酸化,并促进Ⅰ型干扰素的分泌。POctS递送OVA mRNA显著抑制肿瘤生长并诱导免疫记忆,效果优于Alum/OVA疫苗;递送MLKL mRNA可诱导免疫原性细胞死亡,重塑免疫微环境,与PD-L1抑制剂协同抗肿瘤,疗效超越吉西他滨。POctS系统毒性低,炎性因子水平稳定,具有部位特异性累积优势,可精准激活局部免疫而不影响健康组织。
不可降解的PEI存在潜在毒性问题,而可生物降解的聚β-氨基酯(PBAE)凭借易合成、单体易获取等优势成为更优替代选择。Chen等合成了108种结构多样的聚β-氨基酯,通过调控背骨亲疏水性、侧链碳链长度和端基氨基官能团,将其与辅助脂质等结合形成聚合物-脂质纳米颗粒(PLNP),以开发高效靶向递送碱基编辑工具的非病毒载体。该团队筛选出新型聚合物7C8C8,其mRNA递送效率超越FDA批准的脂质载体(如ALC-0315等),且安全性优异(LD50=403.8 mg/kg)。该载体冻存后稳定性长达2个月,体内碱基编辑效率达70%,并能通过协同敲除PCSK9和ANGPTL3基因,实现LDL-C持续降低60%以上。树状大分子作为具有球形分支结构的聚合物,其中聚酰胺胺(PAMAM)是研究最广泛的类型。Tang等设计了一种新型双靶向mRNA纳米颗粒(HDPM NP),其核心由阳离子PAMAMG0-C14包裹mRNA构成,通过表面修饰透明质酸(HA)靶向肺癌细胞表面的CD44受体,同时采用DSPE-PEG-甘露糖(Man)进一步功能化纳米颗粒表面,使其兼具靶向炎症巨噬细胞表面葡萄糖转运蛋白的能力。除合成载体外,壳聚糖等天然聚合物因其良好生物相容性也成为核酸载体候选,常与PLGA、透明质酸等材料联用以提升mRNA稳定性和递送效率。需要指出的是,聚合物材料的批次间差异可能影响药物一致性——分子量的微小变化会显著改变体内mRNA转染效率,这导致尽管聚合物材料种类丰富,其在mRNA治疗领域的临床转化进度仍滞后于脂质系统。
2.4 基于多肽或蛋白质的递送系统
带正电荷的肽和蛋白质可以通过静电相互作用与带负电荷的mRNA结合,从而实现良好的生物相容性和可降解性。两亲性肽因其含有精氨酸等阳离子或两亲性氨基,可通过静电作用与mRNA结合,并借助质子海绵效应促进溶酶体逃逸,从而协助mRNA递送入细胞。Qiu等报道了一种新型RNA递送载体PEG12KL4,该载体通过将合成的KL4阳离子肽与12聚体单分散线性PEG共价连接而成。当PEG12KL4与mRNA以10∶1(质量比)比例复合时,可形成纳米级复合物,并在人肺上皮细胞中实现高效转染。
在蛋白质类递送系统中,鱼精蛋白作为一种富含精氨酸的小分子带正电荷蛋白,能够有效包载mRNA,并已应用于mRNA治疗领域。CureVac公司开发的RNAactive®平台即采用了游离mRNA与鱼精蛋白复合mRNA的组合,可同时诱导平衡的适应性免疫应答和T细胞介导的免疫反应。基于该技术,CureVac已启动多个临床研究项目。
近年来,VLP技术备受关注——这类颗粒仅包含病毒组装衣壳所需的主要结构蛋白,而不携带病毒基因组物质。VLP既保留了病毒载体的高效递送和靶向能力,又避免了病毒基因组整合风险,被用作mRNA疫苗的递送载体。VLP凭借与天然病毒相似的结构,能高效被APC识别和摄取,并模拟病毒感染有效递送mRNA,且无基因组整合风险;其优势包括高生物相容性、固有佐剂效应和优异靶向性,但也面临生产工艺复杂、预存免疫干扰、载量有限及免疫原性难以控制等挑战。
MS2衣壳与特定的19个核苷酸茎环(pac位点)相互作用,将目标RNA包裹进VLP中。Li等利用此方法构建了针对前列腺癌基于MS2 VLP的 hPAP-GM-CSF mRNA 疫苗。此mRNA疫苗引发了强烈的体液和细胞免疫反应,特别是抗原特异性细胞毒性T淋巴细胞(cytotoxic T lymphocytes,CTL)和平衡的Th1/Th2反应,并完全保护C57BL/6小鼠免受前列腺癌的侵害。随后,该团队利用此方法将MicroRNA-122包裹进展示HIV TAT肽的MS2 VLP中用于治疗肝细胞癌(hepatocellular carcinoma,HCC)。结果表明,展示TAT的MS2 VLP能够有效穿透细胞膜并递送miR-122。此外,其在Hep3B、HepG2和Huh7细胞及Hep3B相关动物模型中的对HCC的抑制效果显著。Unti等利用MS2衣壳蛋白将含MS2茎环结构的CVB3 IRES-circRNA包装至VLP,相较于线性mRNA显著增强蛋白表达持久性,但需哺乳细胞体外转录的特性限制了低成本大规模生产。Su等利用双启动子系统同步驱动MS2衣壳蛋白表达与OVA mRNA转录,并通过pac序列介导的特异性识别,实现mRNA在VLP组装过程中的高效包装,获得MS2-OVA mRNA疫苗。基于VLP系统可有效保护mRNA免受核酸酶降解,同时确保稳定性和向宿主细胞的高效递送。
豇豆褪绿斑驳病毒(cowpea chlorotic mottle virus,CCMV)衣壳可包裹2500~4200 nt的异源单链RNA,研究者利用该平台递送连接诺达姆拉昆虫病毒(Nodamura virus,NoV)RdRP的卵清蛋白表位SIINFEKL。未成熟树突细胞与这类VLP共孵育后,成熟标志物激活增强,RNA摄取/复制水平提升,并显著增加抗原特异性T细胞。针对COVID-19的VLPCOV-02疫苗在VLPCOV-01(含甲病毒非结构蛋白1-4和膜锚定RBD的LNP-saRNA平台)基础上进行了改进:引入HLA DR通用结合表位序列(可结合常见人类白细胞抗原DR亚型)和5-甲基胞苷。这些修饰在Ⅰ期临床试验中降低了反应原性和不良事件发生率,同时诱导了强效抗体应答。
在多项研究中,HIV的env和gag蛋白被用于构建基于gag的VLP以递送mRNA。Zhang等将SARS-CoV-2刺突蛋白的天然胞质尾(cytoplasmic tail,CT)与HIV-1或猴-人免疫缺陷病毒(SIV)的gp41跨膜env蛋白CT融合,形成杂交刺突蛋白,其中截短至745个氨基酸的反转录病毒CT能诱导刺突蛋白在细胞表面表达。通过以1∶2比例共转染表达杂交刺突蛋白的mRNA与SIV gag mRNA可形成VLP,而共表达截短刺突蛋白与SIV Gag的mRNA疫苗能激发更高效的免疫应答,其机制可能与多重抗原呈递、膜锚定蛋白天然构象保留或多抗原协同作用有关。另一项研究表明,乙肝病毒(hepatitis B virus,HBV)核心蛋白(hepatitis B virus core protein,HBc)C端截短是决定RNA降解的关键因素,其自组装域(aa 1~140)和鱼精蛋白样聚精氨酸域(PA,aa 150~183)中,去除PA域可减少RNA降解,但保留全部四个精氨酸模块(block1-162、block2-163、block3-171、block4-175)能轻微提升包装效率,而包装能力缺失会促使Th2免疫应答转为Th1型并降低IFN-γ水平,这可能与CTL表位丢失有关,这些发现提示蛋白C端截短是开发高效疫苗的有力工具。类似策略在抗猴-人免疫缺陷病毒mRNA疫苗中同样有效,研究表明在HEK 293T细胞中共表达HIV-1 env与SIV Gag蛋白的mRNA可形成VLP,诱导广谱中和抗体并降低恒河猴感染风险,表明Gag基VLP-mRNA疫苗设计灵活且对多种传染病有效。此外,人类基因组编码的类逆转录病毒gag蛋白(如PNMA家族成员PNMA2)可自组装为二十面体衣壳,经改造后能携带核酸,展示了其在哺乳动物细胞中作为递送工具的潜力。
总结与展望
mRNA技术的崛起无疑是生物医学领域的一场革命。其在COVID-19疫苗中的成功应用,不仅证明了该技术在应对突发公共卫生事件中具有无与伦比的开发速度与防护效力,更展现出作为平台型技术在肿瘤治疗、遗传性疾病防治、蛋白替代疗法等诸多领域的广阔前景。当前,mRNA技术的核心发展方向聚焦于三大维度:分子结构的创新设计(如环状RNA、自扩增RNA等新型核酸平台)、递送系统的持续优化(包括LNP组分工程化改造与新型载体开发)以及生产工艺的协同升级,旨在系统性平衡并提升疫苗的免疫原性、表达效力、生物安全性及制剂稳定性。
值得注意的是,mRNA疫苗技术在兽医学领域同样展现出显著的应用潜力。与传统兽用疫苗相比,mRNA疫苗具备研发周期短、易于针对病原变异进行快速序列更新、可灵活实现多价配伍、通常无需额外佐剂即可激发强免疫应答等突出优势,特别适用于高效应对禽流感、非洲猪瘟、犬瘟热等重要动物传染病的暴发。研究显示,基于LNP或病毒样颗粒(VLP)递送系统的mRNA疫苗在猪、鸡、牛等经济动物中均可诱导强大的体液与细胞免疫,且安全性表现良好。目前已有包括RPS1903在内的多个兽用mRNA疫苗产品获批进入临床研究阶段。随着制剂稳定性技术的突破(如冻干工艺的应用)和大规模生产成本的下探,mRNA技术有望逐步取代传统疫苗平台,成为动物疫病防控的主流技术手段,从而有力推动“人—动物—环境”健康共同体的构建与实践“同一个健康”(one health)战略理念。
然而,mRNA技术迈向更广泛的应用仍面临多项根本性挑战。其一在于制剂的长期稳定性问题,多数现有mRNA疫苗(尤其是LNP制剂)仍严重依赖低温冷链运输与储存,这在资源有限地区可及性较差;其二,递送系统的靶向精度仍有待提高,脱靶效应可能引起安全性顾虑并降低治疗指数;其三,个性化疫苗(如肿瘤新抗原疫苗)的生产成本居高不下,限制了其临床推广与可及性;其四,我们对复杂疾病环境中mRNA表达调控、免疫应答持久性及与宿主互作的深层机制理解尚不全面,制约了理性设计能力的提升。
展望未来,mRNA技术的进一步突破必将依赖于多学科前沿科技的交叉融合。在序列与载体设计层面,可借助人工智能与机器学习方法对mRNA二级结构、密码子使用、调控元件[如UTR、poly(A)尾]进行理性设计,以最大化翻译效率并最小化免疫原性;在制剂开发方面,通过新型冻干工艺、糖基化稳定策略及常温储存型纳米颗粒的开发,有望彻底解决冷链依赖问题;在生产制造领域,模块化、封闭自动化的一体化生产平台将大幅提高产线柔性、降低生产成本,从而推动个体化医疗产品的商业化与应用普及。
最终,随着这些技术瓶颈的逐步攻克,mRNA平台将不再仅限于传染性疾病的预防,而是演进为一个真正可编程的医学工具,能够实现个性化癌症疫苗、蛋白替代疗法、基因编辑工具递送、细胞重编程等多元应用,全面推动人类与动物医学迈向精准、高效、安全的“同一个健康”新纪元。