

细胞膜是生命的边界,一旦遭到破坏往往意味着细胞死亡。穿孔素打孔、Gasdermin 成孔、NINJ1 溶膜 —— 这些经典机制已被深入研究。但细胞膜是否还能以其他的方式被破坏?仍然是结构生物学领域非常重要的科学问题。
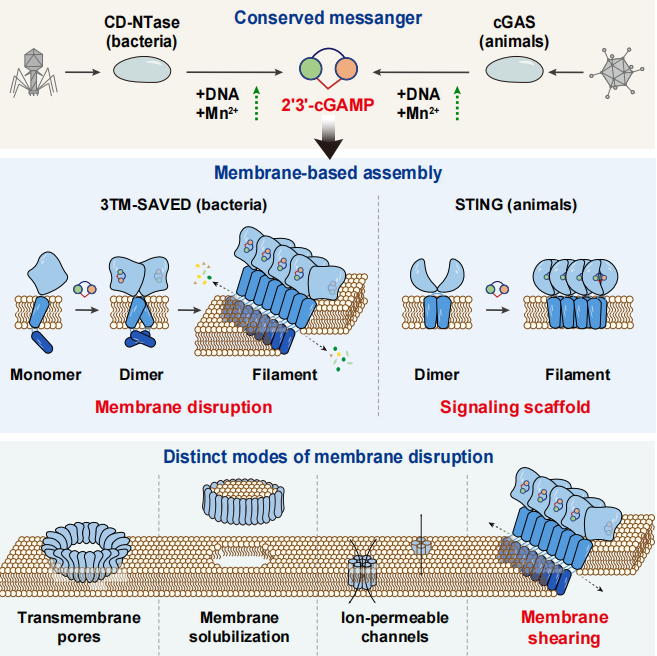
2026 年 4 月 17 日,中国科学院生物物理研究所高璞研究员和北京理工大学高昂教授研究团队在 Cell 在线发表题为「2'3'-cGAMP-induced membrane shearing promotes broad antiphage immunity」的研究论文,揭示了一种全新的膜破坏方式 ——CBASS 跨膜效应蛋白的完整激活与膜破坏机制,提出了 2'3'-cGAMP → 丝状组装 → 垂直脂质剪切 → 膜通透性增强 → 细胞死亡这一信号传导链条,不仅增加了对膜生物学的认知,更为理解细菌到人类的先天免疫进化提供了新线索。
细菌的「自杀式」抗病毒防线:CBASS 系统
按直觉来看,细菌没有复杂的免疫系统,面对噬菌体入侵只能束手就擒。但研究发现,细菌和古菌中广泛存在一种名为环状寡核苷酸的抗噬菌体信号系统(Cyclic-oligonucleotide-based anti-phage signaling system,CBASS)的抗病毒防御机制,与哺乳动物的 cGAS-STING 先天免疫通路在进化上同源。当噬菌体感染时,CBASS 系统的上游酶会合成环核苷酸信号分子,下游效应蛋白感知信号后触发细胞死亡,以「自杀」方式阻止噬菌体复制。然而,超过四成的 CBASS 效应蛋白并非可溶性酶,而是嵌入细胞膜的跨膜蛋白 —— 它们究竟如何识别信号并破坏膜结构,长期困扰着该领域。
从单体到纤维:2'3'-cGAMP 触发的逐级组装过程
研究团队选择苏云金芽孢杆菌(Bacillus thuringiensis)中的一个代表性跨膜效应蛋白 3TM-SAVED(含三个跨膜螺旋和 SAVED 结构域)展开深入解析。令人惊讶的是,这个细菌蛋白的上游酶合成的信号分子竟然是 2'3'-环 GMP-AMP(2'3'-cyclic GMP-AMP,2'3'-cGAMP)—— 与哺乳动物 cGAS 酶产生的分子相同,且同样需要 DNA 和锰离子激活。这种跨越物种的功能保守性暗示,先天免疫的核心机制在进化上远比想象的古老。
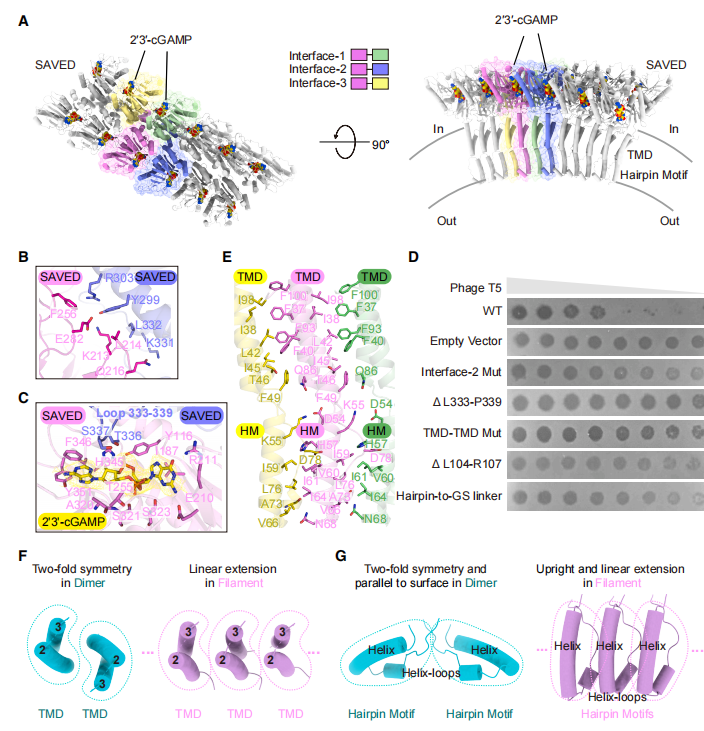
为了理解 3TM-SAVED 如何响应 2'3'-cGAMP,研究团队利用冷冻电镜捕捉了激活过程的动态结构。结果发现,在没有配体时,蛋白以单体形式稳定存在;在加入 2'3'-cGAMP 后,蛋白首先组装成瞬时的二聚体 —— 两个蛋白分子通过 SAVED 结构域和跨膜结构域相互作用,形成一个对称的「花形」结构。但这个二聚体并没有立即破坏膜,而是作为一个中间态等待进一步激活。
随着时间推移,二聚体继续组装成更高阶的纤维状结构。在这种纤维中,蛋白分子呈拱形排列,SAVED 结构域向外凸出,跨膜螺旋形成中央层,而原本平行于膜表面的两亲性发夹基团则转变为直立构象。正是这一构象转换,催生了一种前所未见的膜破坏现象:纤维将脂质双分子层沿垂直方向「剪开」,形成两层错位的膜片。
第三种破膜方式:垂直剪切,而非打孔或溶膜
这种垂直膜剪切的机制与经典的膜破坏方式截然不同。比如,穿孔素和 Gasdermin 通过在膜上打孔造成损伤,NINJ1 通过溶解膜释放膜盘,离子通道则形成选择性通透的孔道。然而,3TM-SAVED 纤维既不形成跨膜孔,也不溶解膜,而是像一把垂直于膜平面的「剪刀」,将双分子层剪成两层并横向错位。被剪切的膜片之间由柔性环连接,产生了线性排列的孔隙,允许水分子、离子和小分子自由通过,最终导致膜通透化和细胞死亡。
接下来,研究团队在苏云金芽孢杆菌中验证了这种机制的真实性:外源添加 2'3'-cGAMP 可诱导表达 3TM-SAVED 的细胞死亡,而不表达该蛋白的对照细胞不受影响;冷冻电镜断层成像显示,处理后细胞的内膜明显弯曲,膜-肽聚糖层分离,丝状密度沿弯曲膜富集,与体外实验观察到的纤维组装高度一致。
有意思的是,生物信息学分析显示,含有类似结构模块的效应蛋白不仅广泛分布于 CBASS 系统,还出现在 CRISPR-Cas 等其他抗病毒基因座中,跨越 Pseudomonadota、Bacillota、Actinomycetota 等多个细菌门。尽管这些同源蛋白的两亲性发夹基团呈现结构多样性 —— 包括本研究发现的螺旋-转角-螺旋/环、β-发夹和环-转角-环等变体,但其双亲性特征高度保守,暗示垂直膜剪切可能是一种被广泛采用的抗病毒策略。
因此,这项工作完整描绘了 CBASS 跨膜效应蛋白从信号感知到膜破坏的分子机制,打破了膜破坏只有「打孔」和「溶膜」两种模式的固有认知,建立了垂直膜剪切作为一种与经典成孔、膜溶解、离子通道不同的全新膜破坏机制。同时,研究证实细菌 CBASS 与哺乳动物 cGAS-STING 在配体识别上的进化保守性,为理解先天免疫的起源和演化提供了重要线索。
不过,垂直膜剪切机制在不同细菌物种、不同膜组成环境中的普适性仍需更多实验验证;当前对广泛保守性的推断主要基于生物信息学分析,有待在更多生物体中进行系统确认。此外,3TM-SAVED 的可控膜破坏特性在合成生物学或抗菌应用中的潜力虽令人期待,但其可行性、可控性和安全性仍需深入评估。